



























肿瘤是一个复杂的生态系统,其中癌症细胞和多种宿主细胞之间的相互作用影响疾病的进展和治疗反应。在肿瘤进展过程中,癌症细胞采用多种途径来逃避免疫攻击。同时,癌细胞劫持免疫细胞,如中性粒细胞、巨噬细胞和调节性T细胞(Treg),以协调免疫抑制性肿瘤微环境(TME)。这反过来促进免疫逃逸,促进血管和细胞外基质的重塑,并支持癌症进展和治疗抵抗。异常免疫反应被广泛认为是癌症的标志,并为癌症治疗提供了有吸引力的靶点。
因此,有必要深入了解癌症细胞的内在特征,包括遗传变异、信号通路调节和代谢改变,它们在协调免疫景观的组成和功能状态方面发挥关键作用,并影响免疫调节策略的治疗效益。

遗传改变
癌基因、抑癌基因或DNA损伤修复基因中的特定癌症细胞固有遗传改变有助于调节癌症免疫状况。
癌基因
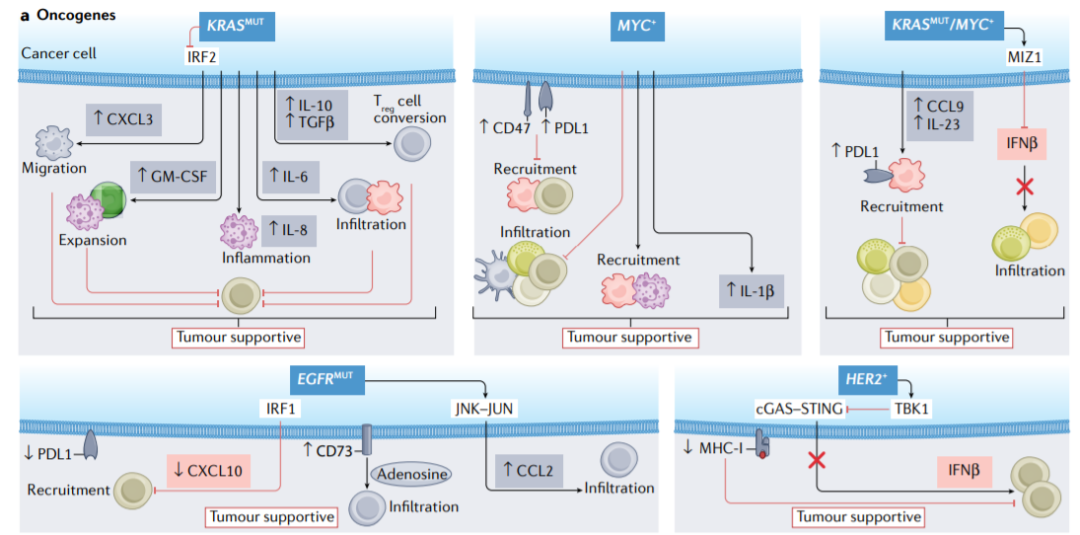
癌细胞中不同的致癌基因通过不同的机制调节免疫行为,这些机制因癌症类型、癌症分期和癌症部位而异。例如,KRAS突变通过IL-8诱导的炎症、IL-6介导的巨噬细胞和Treg细胞浸润、GM-CSF诱导的Gr1+CD11b+髓系细胞的扩增、IL-10和TGF-β介导的CD4+CD25−T细胞向Treg细胞的转化、以及抑制干扰素调节因子2(IRF2)和随后产生的CXCL3,从而导致CD8+T细胞抑制,髓系衍生抑制细胞(MDSCs)的迁移增强。
抑癌基因

抑癌基因通过不同的环境依赖机制调节免疫。例如,研究发现Trp53的缺失通过WNT配体介导的巨噬细胞极化和IL-1β的产生驱动免疫抑制,导致中性粒细胞的全身聚集,从而CD8+T细胞的抑制和CXCL17、CCL3、CCL11、CXCL5和M-CSF介导的巨噬细胞和Treg细胞的募集。
TP53的突变与PD-L1表达相关,并通过CXCL2分泌调节免疫抑制中性粒细胞的募集,通过NF-κB介导的IL-8调节慢性炎症,并且结合TBK1抑制下游cGAS–STING–IRF3信号传导和IFN-β1的产生,其减少T细胞和NK细胞浸润并增强巨噬细胞极化。相反,TP53的功能增益突变与新抗原表达,以及IFN-γ和CXCL9表达相关,这支持抗肿瘤免疫。
DNA损伤修复机制的遗传改变
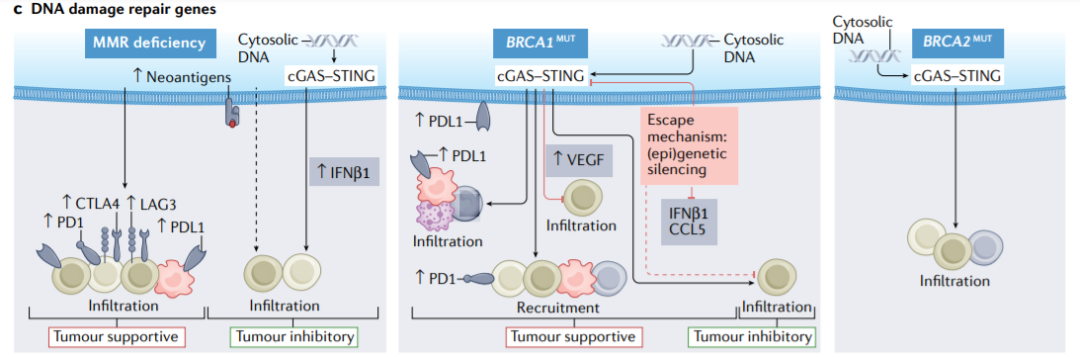
癌症细胞中的错配修复(MMR)缺陷会导致突变、新抗原和细胞中DNA的积累,从而触发cGAS–STING依赖性的T细胞浸润,免疫检查点分子如PD-1、PDL1、CTLA-4和LAG-3在浸润免疫细胞上的表达增强。
在BRCA1突变的肿瘤中,双链DNA积聚在细胞中,并引发与CD8+T细胞浸润相关的cGAS–STING介导的炎症反应。cGAS–STING还促进免疫抑制性髓系细胞、巨噬细胞、Treg细胞和耗竭的PD1+T细胞的募集,以及VEGF的上调。DNA传感基因和下游介质(如IFN-β或CCL5)的遗传缺失或表观遗传沉默可介导BRCA1突变肿瘤的免疫逃逸,以及PD-L1的表达增强。相反,BRCA2突变肿瘤显示cGAS–STING介导的各种T细胞群的富集。
表观遗传修饰
除了遗传改变外,表观遗传改变也是癌症细胞的一个共同特征,在核结构和基因转录中起着至关重要的作用。癌症表观基因组会发生各种变化,包括DNA甲基化、组蛋白修饰、染色质重塑和非编码RNA的失调。越来越多的证据表明,癌症细胞中发生的表观遗传变化也会影响与免疫系统的串扰。
DNA甲基化

由于超级增强子(SE)的形成,炎症基因的DNA去甲基化导致CXC趋化因子配体(CXCL)介导的中性粒细胞募集,而总体低甲基化与PD-L1、IL-6和VEGF的表达增强相关。而DNA(超)甲基化可降低编码PD-L1的基因CD274和编码MHC-I的HLA基因的表达,而编码cGAS–STING的基因通过启动子甲基化的转录抑制增强MHC-I表达和T细胞识别。
组蛋白甲基化
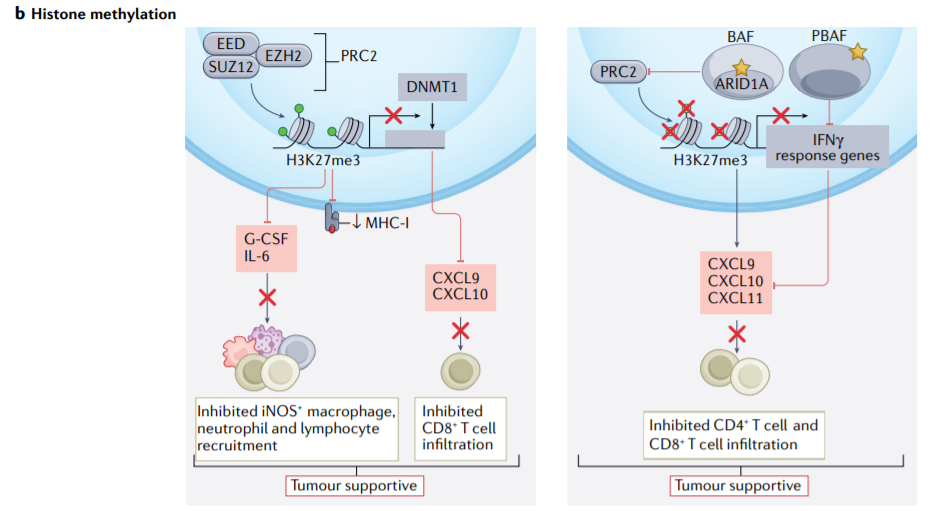
组蛋白H3在Lys27发生三甲基化,在DNA低甲基化的背景下,抑制编码MHC-I、CXCL9和CXCL10的基因,并减少CD8+T细胞浸润。
癌症细胞中的多梳抑制复合物2(PRC2)活性介导H3K27me3,抑制G-CSF和IL-6的表达并诱导性一氧化氮合成酶阳性(iNOS+)巨噬细胞、中性粒细胞和T细胞的招募。ARID1A是SWI/SNF复合物的DNA结合亚基, ARID1A的遗传突变会抑制IFN-γ诱导基因转录和CXCL9、CXCL10和CXCL11的表达,从而抑制T细胞浸润。
组蛋白乙酰化

组蛋白乙酰化通过组蛋白乙酰转移酶1(HAT1)调节免疫应答,HAT1介导CD274和组蛋白去乙酰化酶(HDACs)的表达,HDACs抑制编码PD-L1和PD-L2的基因的表达,以及CCL5、CXCL9和CXCL10的表达,从而损害T细胞浸润。
细胞内信号通路
癌症细胞的一个关键特征是异常信号传导,这是由编码或非编码DNA区域的遗传或表观遗传改变以及生长因子和代谢等外部信号驱动的。
WNT–β-catenin通路
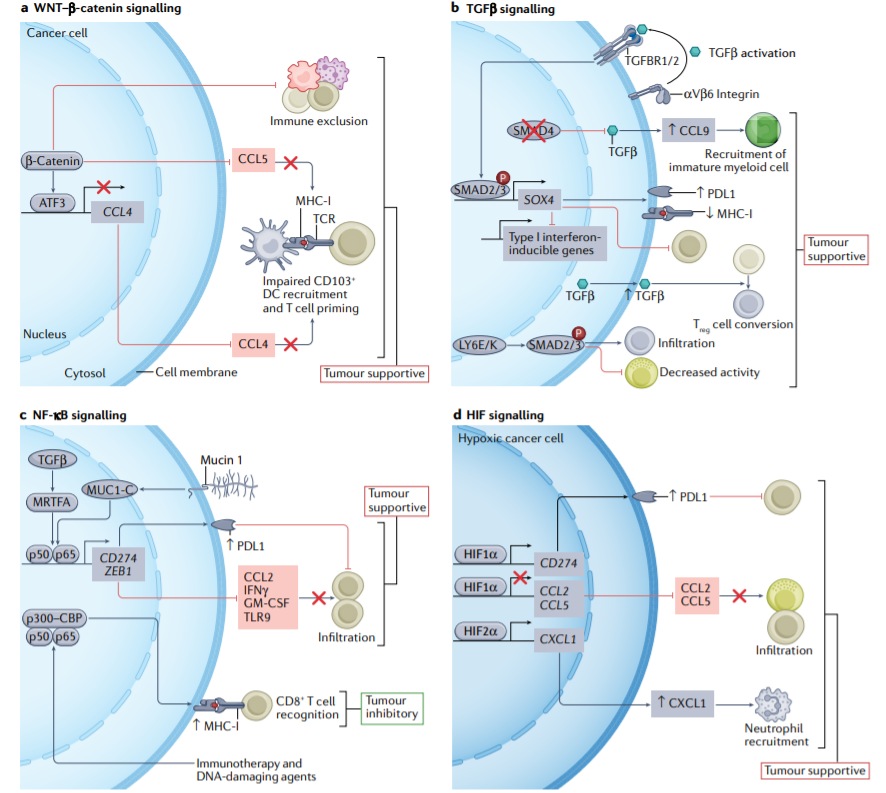
激活的癌症细胞内固有WNT-β-catenin信号与免疫排斥相关,但其潜在机制仍不明确。WNT-β-catenin通过诱导ATF3表达阻断CD103+树突状细胞的募集和T细胞启动,并抑制CCL4或CCL5分泌。
NF-κB信号通路
PD-L1的表达受癌症细胞内NF-κB信号转导的转录调控,其途径是通过MUC1-c与CD274启动子上的NF-κB亚基p65的直接结合,或通过TGF-β介导的MRTFA的表达。MUC1-C和p65还驱动ZEB1转录,这导致编码CCL2、IFN-γ、GM-CSF和TLR9的基因受到抑制,CD8+T细胞浸润受损。相反,免疫疗法和化疗诱导的NF-κB和p300–CBP的活化增强了MHC-I抗原呈递和CD8+T细胞识别。
HIF信号通路
缺氧诱导的缺氧诱导因子1α(HIF1α)信号通路通过增强CD274的表达和抑制编码CCL2和CCL5的基因来驱动免疫逃避,这会损害NK细胞和CD8+T细胞的浸润,而HIF2α诱导CXCL1的表达并促进肿瘤的中性粒细胞募集。
代谢改变
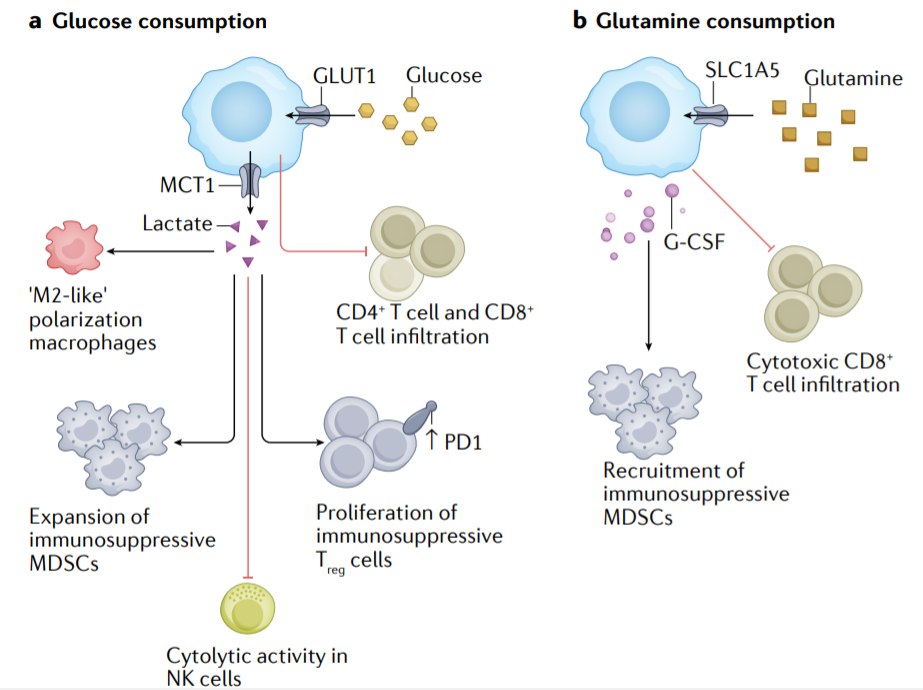
在肿瘤的发生和进展过程中,癌细胞及其TME不断暴露在恶劣的条件和压力下。为了生存和维持生长,需要细胞适应和代谢重编程。癌症代谢重编程与免疫抑制和逃避有关。
肿瘤细胞消耗大量葡萄糖,这与T细胞浸润不良有关。高葡萄糖消耗导致乳酸的产生并分泌到肿瘤微环境中,乳酸在肿瘤微环境中以免疫抑制的方式起作用,降低NK细胞的细胞溶解活性,并增强PD-1表达和Treg细胞的免疫抑制能力。此外,乳酸增加了肿瘤和脾脏中MDSC的频率,并在肿瘤相关巨噬细胞中诱导“M2样”极化。此外,癌症细胞中谷氨酰胺的消耗降低CD8+T细胞的活化和浸润,并通过增加G-CSF的分泌增强MDSC的募集。
近年来,关于癌症细胞-免疫细胞串扰的研究大幅增长。跨越式发展的技术进步,包括单细胞多组学技术,基于人工智能的系统生物学方法以及体内体细胞基因编辑策略,有望加速这些研究的深入,并最终形成针对个体患者的新型免疫干预策略。
参考文献:
1.Mechanisms driving the immunoregulatory function of cancer cells. Nat Rev Cancer.2023 Jan 30.
声明:内容及图片素材整理至网络,版权归原作者所有,本文仅供学习科普,无任何商业用途。如有侵权或错误,请速与管理员联系,我们将及时更正/删除,谢谢。
往期回顾 | |








 浙公网安备33011002015279
浙公网安备33011002015279 本网站未发布麻醉药品、精神药品、医疗用毒性药品、放射性药品、戒毒药品和医疗机构制剂的产品信息
本网站未发布麻醉药品、精神药品、医疗用毒性药品、放射性药品、戒毒药品和医疗机构制剂的产品信息
收藏
登录后参与评论